
间充质干细胞(mesenchymalstem cells),或者,间充质基质细胞(mesenchymal stromal cells),缩写都是MSCs。本文试图以时间轴为主线,讲述MSCs的研究历程上比较重要的事件。
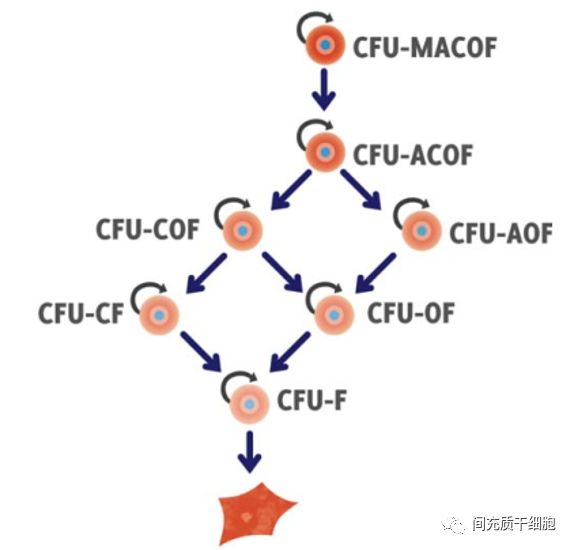
1966年,Friedenstein教授发现骨髓细胞移植能在体内分化为成骨细胞[1],随后在1968年提出存在成骨细胞和造血细胞的祖细胞[2],1971年通过动物实验进一步证实成骨祖细胞的存在[3],1974年成功通过贴壁分离培养的方法, 发现从骨髓中贴壁分离一种成纤维细胞样的细胞,具有分化为成骨细胞的细胞集落(CFU-F),而且支持造血克隆的形成[4, 5];当时还没意识到这种骨髓基质细胞具有多项分化潜能;1987年明确提出这种骨髓基质细胞(成纤维细胞克隆形成细胞,fibroblastcolony-forming cells,FCF-C)为成骨干细胞(osteogenic stem cells),经过20-30次的倍增增殖后,植入大鼠腹膜内依然可以形成骨组织[6];1988年,将这种成骨祖细胞命名为骨髓基质干细胞(marrow stromal stemcells)[7, 8]。
成骨干细胞除了osteogenicstem cells这种写法,还有一词osteoprogenitors,其最早见于1984年关于细胞核大小的描述[9]。1990年确定osteoprogenitors属于基质细胞(mesenchymal cells),具有分化成骨细胞和软骨细胞的能力[10]。
1987年,Charbord博士发现在马血清的培养下,骨髓基质细胞在摄取血清蛋白方面有别于骨髓成纤维细胞,从而明确了MSCs并不是成纤维细胞,虽然在体外培养时的细胞形态和成纤维细胞很相似[11]。
1991年北爱尔兰Queen's University Belfast的一个实验室从脐带华通氏胶(Wharton'sjelly)中通过组织块培养的方法,培养出MSCs,当时称呼为“成纤维细胞样细胞”,至少能传代至第6代,能分泌胶原蛋白和细胞角蛋白(CK)等[12]。但是当时对MSCs的研究和认识都不深刻,尚未认识到MSCs的强大免疫抑制和分泌细胞因子的功能。2000年,智利大学的一个实验室从脐血中分离出单个核细胞,然后贴壁纯化培养获得MSCs[13]。随后,不同的实验室在不同的组织中均分离和培养出MSCs,比如脂肪、羊膜、牙龈、胸腺、胎盘、滑膜、胎血、胎肝、胎肺等。
1991年Charbord博士开始用一些细胞表面标记物来识别MSCs:CD45、HLA-DR 、CD33 、CD11c、vW因子、a-SMA、纤维连接蛋白(fibronectin)、CD68、CD51[14]。1993年Charbord博士检测了MSCs细胞表型:CD10、CD14、CD34、CD45、CD44、LFA-1α、LFA-1β、LFA-3、CD29、VLA-1、VLA-2、VLA-4、VLA5、VLA-6、Stro-1、VCAM-1[15]。
1992年,来自于Friedenstein教授团队的Owen博士证明MSCs同时具有向脂肪细胞和成骨细胞分化的能力;培养基中存在地塞米松的情况下,MSCs向脂肪分化占主导地位,抑制了向成骨细胞分化;在地塞米松和1,25-二羟维生素D3共同培养时,脂肪细胞的分化受到抑制,而成骨细胞的分化则增强[16]。1999年《science》首次证明MSCs在体外同时具有向脂肪细胞、成骨细胞、软骨细胞分化的能力[17]。真是这篇文章,点燃了大家对MSCs的研究热情,集中在研究MSCs的各种分化功能,期待着MSCs在再生医学中发挥巨大的作用。
虽然1974年Friedenstein教授认识到MSCs参与了造血微环境的一个组成细胞支持造血[5],但到1992年才明确提出MSCs作为滋养层细胞支持造血干细胞向粒细胞、巨噬细胞和巨核细胞分化[18, 19],而且这种支持作用在于MSCs分泌这些生长因子(CSF-1、GM-CSF、G-CSF、IL-6、c-kitligand和IL-3)[19]。
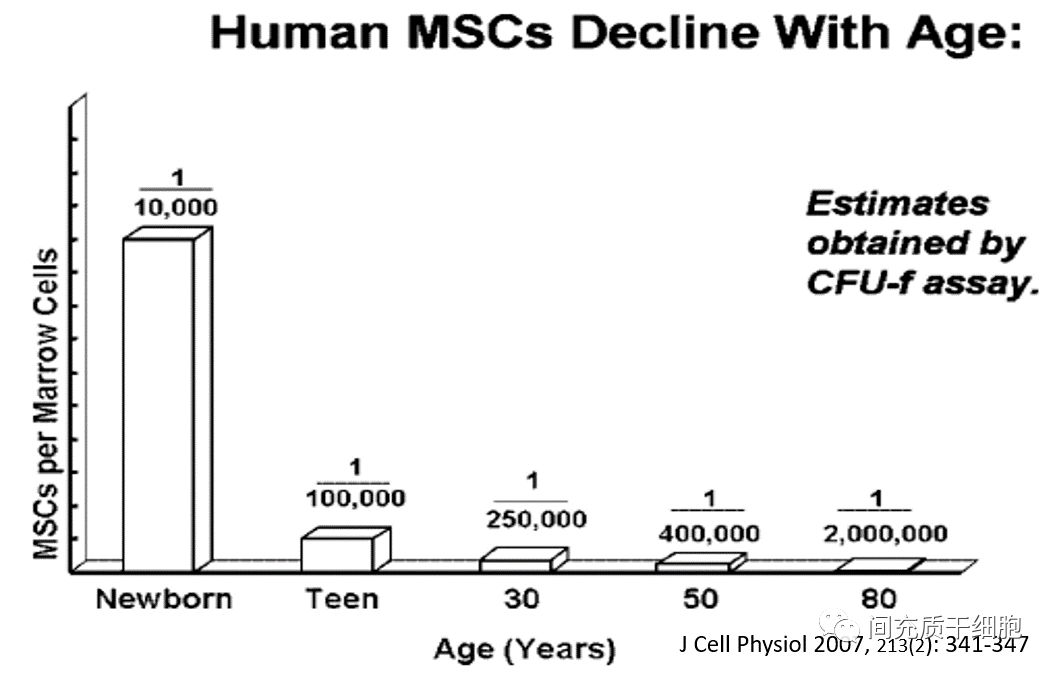
1992年发现年龄大的大鼠骨髓MSCs的数量较年轻者少,即成纤维细胞集落形成单位(FCF-U)少,贴壁细胞少[20]。1999年发现在人体上存在同样的现象,即骨髓MSCs的数量随着年龄增大而减少[21]。
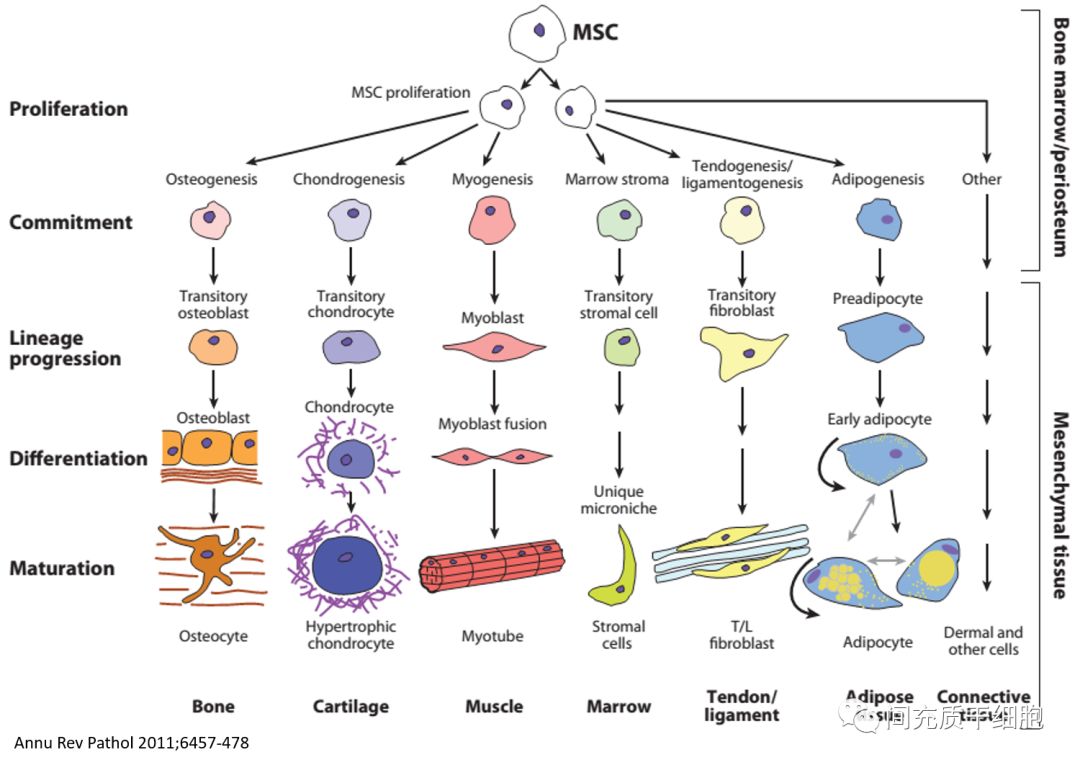
不同于Friedenstein教授和Charbord博士是研究人的骨髓MSCs,Caplan教授刚开始是研究鸡胚胎的骨髓MSCs,根据鸡胚胎骨髓MSCs的特性,提出MSCs是来源于中胚层(Mesodermal),结合前人的研究成果,于是在1991年将MSCs命名为间充质干细胞(Mesenchymal Stem Cells,MSCs)[22]。谁都没想到,MSCs被命名为间充质干细胞后,这个名称备受欢迎,大量的研究文章都采用这个名字。下面的一张MSCs体外分化示意图,至少出现在Caplan教授的四篇文章中(Tissue Eng Part A, 2010/Annu Rev Pathol, 2011/Cell Stem Cell, 2011/StemCells Transl Med, 2017)。
1994年Caplan教授连发3篇文章提出自体MSCs在骨和软骨的组织再生医学方面有非常好的治疗前景[23-25]。1995年报道MSCs的第一篇临床研究成果,从恶性血液病患者骨髓抽取并分离培养出这些贴壁的基质细胞,然后回输到患者体内,观察临床效果并证明这些基质细胞的安全性[26]。这是一个足以载入史册的研究,是间充质干细胞研究史上的第一个里程碑式的事件,使间充质干细胞的研究从实验室跨入到实际的临床应用。在1990年代,Caplan教授和他的同事创办的Osiris治疗公司,生产制备MSCs注射液(Prochymal)于2012年在加拿大有条件获批,用于儿童移植物抗宿主病(GVHD)的治疗。
1997年Darwin J. Prockop博士在《science》撰文,提出MSCs为骨髓基质细胞(Marrow Stromal Cells)[27];随后在1998年提出MSCs可以作为基因治疗的载体[28]。2002年提出根据细胞的大小来判断MSCs的质量,即培养过程中,胞体小的MSCs具有最强的成脂潜能与产生单细胞集落的能力[29]。
2000年《nature medicine》采用人骨髓MSCs子宫内注射到孕65天和85天的羊胎(相当于145天的人胎)的腹膜腔,移植后长达13个月的时间内在多个组织中存活,而且移植后的人MSCs在体内分化为软骨细胞、脂肪细胞、心肌细胞和心肌细胞、骨髓基质细胞和胸腺基质细胞;这研究结果直接证明了MSCs的多项分化潜能[30]。
2000年,一个关于细胞治疗的学术会议上,Charbord博士提出CD34/45阴性、CD10/13阳性和Thy-1(CD90)阳性是MSCs表型的一个关键成分;Simmons博士建议在骨髓收获后立即研究间充质细胞,以Stro-1阳性,糖激素A阴性作为鉴定这些细胞的标准,并建议将STRO-1、Thy-1、VCAM-1阳性细胞和CD45阴性细胞作为候选群体;研讨会建议,鉴定间充质细胞的最小决定因素可能是Thy-1、Stro-1、平滑肌肌动蛋白、I型胶原和vimentin,而CD45阴性,尽管在这一点上没有明确的共识[31]。Charbord博士和Simmons博士是最早关注MSCs的细胞表面标记物的科学家。
2002年Charbord博士和Caplan教授联合研究证明Stro-1阳性MSCs为多潜能干细胞[32],但是2004年Charbord博士参与的研究团队发现Stro-1阳性MSCs和Stro-1阴性的MSCs均有归巢的功能,而Stro-1阳性MSCs却不能支持造血[33]。
2002年,2个团队分别发现了间充质干细胞有强大的免疫抑制能力[34, 35]。2003年Le Blanc博士发现MSCs本身具有低免疫原性,即使异体或跨种属使用,均不会引起免疫排斥反应[36]。LeBlanc博士在2004年《Lancet》发表MSCs治疗移植物抗宿主病(GVHD)的临床研究的文章[37]。这是第一篇利用MSCs的免疫抑制能力来发挥治疗作用的临床研究文章。广泛性免疫抑制功能可以说是MSCs专属的独特性功能,对多种免疫细胞均有抑制活化的功能。具有类似功能的Treg细胞,抑制范围仅限于T淋巴细胞。MSCs的免疫抑制功能的发现,也极大地推动了MSCs的临床研究,用于治疗免疫反应性疾病。
2004年Paolo Bianco教授建议MSCs命名为骨骼干细胞(skeletal stem cells),而且通过转基因小鼠证明持续升高的甲状旁腺激素通过甲状旁腺激素相关肽受体,促进MSCs向骨细胞分化,成骨祖细胞增多,但是同时耗竭骨髓中的MSCs[38]。PaoloBianco教授随后根据发育学的一些相关证据,对骨骼干细胞的本质进行多方面的阐述,指出MSCs的本质就是骨骼干细胞,强调骨骼干细胞本身的正确概念对于正确的临床应用有密切相关性[39-41]。
2005年国际细胞治疗协会(民间组织)认为MSCs在体内没有分化能力,不具有“干细胞”的特性,建议MSCs命名为多潜能间充质基质细胞(multipotent mesenchymal stromal cells)[42],并在2006年提出多潜能间充质基质细胞的最低定义标准:①贴壁培养,②特异性表面抗原表达(细胞表型),③向成脂成骨和软骨细胞分化的能力[43]。本小编对这个命名表示颇为不理解,既然认定MSCs为多潜能,那就意味着MSCs有多项分化能力;有了多项分化能力,为何不可以称为“干细胞”?而且那么多的动物实验都证明MSCs可以在体内分化为成骨细胞和软骨细胞,难道这都还不能称为“干细胞”?大咖也会为了推出自己的观点而选择性忽视对自己不利的证据。由于尚未找到MSCs特异性细胞表面标记物,研究者多参考国际细胞治疗协会提出的定义标准。
2005年首次发现病人(再障)的骨髓MSCs的免疫抑制功能出现病理性降低[44],但不清楚疾病导致MSCs的免疫抑制功能降低,还是先出现MSCs的免疫抑制功能降低再出现疾病?随后发现多发性骨髓瘤、系统性红斑狼疮、类风湿性关节炎和ITP等患者的骨髓间充质干细胞的免疫抑制能力也出现损伤,以及增殖速度减慢、分泌细胞因子减少等改变[45]。
1990年Diaz-Flores博士发现骨骼和骨骼周围组织血管周围有一种细胞具有祖细胞的特性,能促进血管新生和组织修复的功能,鉴定为血管周细胞(pericytes)[46];随后2年内证明这种周细胞具有向成骨细胞和软骨细胞分化的能力[47, 48]。1993年Bianco博士利用激光共聚焦技术观察到骨髓基质细胞定位在骨髓血管周围[49]。2006年小鼠实验证明MSCs分布在全身血管周围,因此几乎全身所有的器官和组织基质中均存在MSCs[50]。2008年Bruno Peault博士的研究清楚地证明,从各种组织中分离出来的周细胞产生的MSCs,都具有相同的细胞表面抗原及其体外多能性[51]。同年Caplan教授提出大多数间充质干细胞是从血管周围或壁细胞(周细胞)分化而来的[52]。重要的是,MSCs可以从每个血管化的组织中分离出来[53],甚至可以从月经中分离出来(即破碎的血管释放血管周围的细胞,并分化为MSCs)[54, 55]。脂肪组织的周细胞和MSCs具有相似的基因谱[56]。MSCs来源的血管周围位置及其免疫调节和营养功能(包括制造和分泌抗生素蛋白[57])对MSCs的“基质”名称和来源提出了挑战。2017年中美科学家使用可诱导的TBX18-CreERT2系进行的系谱追踪实验表明,周细胞和血管平滑肌细胞在衰老和不同的病理环境中保持着它们的特性,和MSCs有着不同的行为特征,比如在衰老和疾病状态下MSCs具有成脂分化[58]。
那么,MSCs到底是不是周细胞?如果是的话,那就能解释为何在身体的多个组织器官均能发现MSCs的存在,因为有血管的存在,那就有周细胞的存在。如果不是,那么MSCs和周细胞在发育胚胎学上有什么关系?
2009年,有学者认为人脐带的血管周围细胞(umbilical cord perivascular cells)属于脐带MSCs的一个亚群,具有MSCs相似的功能,提出MSCs群体的异质性,即不同的亚群之间的分化功能略有区别,MSCs随着培养时间延长,多项分化功能逐渐丧失,而均一化为“成纤维细胞”[59](见下图)。2012年和2017年的两篇综述文章对这个观点表示认可[60, 61],但是都没有实锤证据。如果MSCs经过长期培养后变成成纤维细胞,即丧失向脂肪细胞、成骨细胞和软骨细胞分化的能力。实际上,长期培养后,脐带MSCs(P30)依然具有向脂肪细胞和成骨细胞的能力[62]。如果根据MSCs的细胞大小来断定MSCs的异质性,那也是不科学的,因为MSCs在培养过程中,处于不同的细胞分离周期,MSCs的胞体大小当然不一样。关于MSCs异质性可能最直接的证据就是:几乎所有的MSCs均能分化为成骨细胞,但不是所有的MSCs都能分化为脂肪细胞。那么,如何理解MSCs异质性?
在上个世纪的70年代,MSCs的体内分化能力已经得到确认。而MSCs体内自我更新的能力直到2007年和2010年的两篇关于骨髓niche的研究才得到体内证据的支持[63, 64],当在体内移植时,这些细胞产生一个具有合适的组织学和结构的微环境(niche),在这个niche中,接受造血干细胞移植的动物的造血功能得以建立。即MSCs是能够在体内产生一个完整的异位骨或骨髓器官(Ossicle)的单个细胞,具有与原先移植细胞相似的表型和特性的血管周围基质细胞[63]。骨骼组织的祖细胞(MSCs)既是造血微环境的组织者和造血功能维护者[63-65]。PaoloBianco教授认为这种在体内的潜力是系统限制的和细胞自主的,并且不需要与骨形态发生蛋白(BMPs)相结合,MSCs在体内不具有产生非骨骼组织细胞的潜能[66]。所以,Paolo Bianco教授建议MSCs命名为骨骼干细胞。但是,把MSCs注射到羊的胚胎中,MSCs却可以分化为软骨细胞、脂肪细胞、心肌细胞和心肌细胞、骨髓基质细胞和胸腺基质细胞[30]。因此,即使是大咖,也会为了支持自己的观点而选择性地忽略掉一些对自己不利的证据。
2008年,《Nat Rev Immunol》提出MSCs的作用方式为“Touch and Go”(接触后离开),即MSCs通过其快速迁移到受损的器官,并释放应激诱导的治疗分子后被机体清除[67]。2010年另一篇文章提出类似的观点,即“Hit and Run”(打击后离开)[68]。
2010年,Caplan教授建议将MSCs的名称改为药物信号细胞(Medicinal Signaling Cells),名字的缩写也是MSCs[69],以更准确地反映这些细胞位于损伤或疾病部位并分泌生物活性因子[70]。这些生物活性因子具有免疫调节和营养的功能[71],这意味着这些细胞制造药用的治疗物质[72]。实际上,正是患者自身的部位特异性和组织特异性的干细胞构成了新的组织,而这些新的组织是由外来MSCs分泌的生物活性因子刺激组织自身干细胞所产生的[73, 74]。2017年Caplan教授认为MSCs属于周细胞,并提出新的假设:MSCs作为黑色素瘤向骨转移的守护者[75]。2018年,有专家认为将MSCs重新归类为“基质细胞”或“信号细胞”无助于解决MSCs的最终本质和生物学功能特性[76]。无论称呼如何,指导其临床开发和使用的科学原则必须与其他新疗法的原则相同:精确性、有效性、定性、客观性,而不是具有误导倾向的术语和概念模型[76]。毫无疑问,2019年,Caplan教授针对这篇文章给予回应,而且继续呼吁MSCs并非干细胞,而是药品信号细胞[77]。命名MSCs为干细胞的,是你;呼吁MSCs不是干细胞的,也是你。
2011年7月,韩国食品药品管理局准许由FCB-Pharmicell公司开发的MSCs药物Hearticellgram-AMI在韩国上市,适应症为急性心肌梗塞。Hearticellgram-AMI成为全球首个利用自身骨髓MSCs制成的治疗急性心肌梗死的干细胞药物,给药途径为局部冠状动脉注射。2012年1月中旬连续批准Medi-post公司的软骨再生治疗药物Cartistem和Anterogen公司的肛瘘治疗药物Cuepistem的生产许可。Medi-post公司的Cartistem是从脐带血中分离培养的MSCs专门治疗退行性关节炎和膝关节软骨损伤的干细胞药物。Anterogen公司的Cuepistem是从患者本人的脂肪组织中分离培养的MSCs治疗复杂性Crohn’s病并发肛瘘。
2012年,Osiris公司宣布全球首个MSCs的III期临床试验失败,即MSCs治疗儿童GVHD的III期临床试验的结果和安慰剂组没有差异,给与MSCs行业一个几乎致命的打击。当时的III期临床试验的数据并没有整理发表文章,但是2013年有篇文章专门点评这件事,分析失败原因和今后的发展方向[78]。当然,Prochymal 失败的原因有很多,本小编也曾详细地分析过[79]。Prochymal(MSCs)的失败,促进了对MSCs本身进行深入的研究,比如MSCs的分离技术、培养技术、MSCs细胞质量、群体异质性、供体的健康状态等等方面。同时,2012年美加两国研究者发现液氮冷冻运输至病床边复苏的做法对MSCs的免疫功能有损害[80]。虽然美国FDA没有批准Prochymal上市,但是Prochymal却在加拿大获得有条件上市。随后,英国(2012年)和欧盟(2014年)的治疗指南推荐MSCs作为治疗2-4级急性GVHD的三线治疗药物[81, 82]。
2016年7月份《Lancet》报道全球第二个MSCs的III期临床试验的结果,即Cx601(脂肪MSCs)治疗复杂肛周瘘克隆恩病的多中心随机对照III期试验(RCT),这项研究包括212名患者,107名患者接受了ASCs治疗,105名患者接受了安慰剂治疗;接受ASCs治疗的患者中有50%实现了联合缓解,而安慰剂组中的34%的患者在24周达到了联合缓解,MSCs治疗的效果明显优于安慰剂组[83]。Cx601治疗复杂肛周瘘克隆恩病的长期疗效观察结果在2018年发表,在52周时,接受Cx601治疗的患者中达到联合缓解的比例(56.3%)显著高于对照组(38.6%)(P=0.010),临床缓解率(59.2%vs 41.6%,P=0.013);Cx601组不良事件发生率为76.7%,对照组为72.5%[84]。这次III期临床试验的结果明显优于对照组,欧盟在2018年3月批准Cx601(Alofisel)上市用于克罗恩病患者复杂性肛周瘘的治疗。

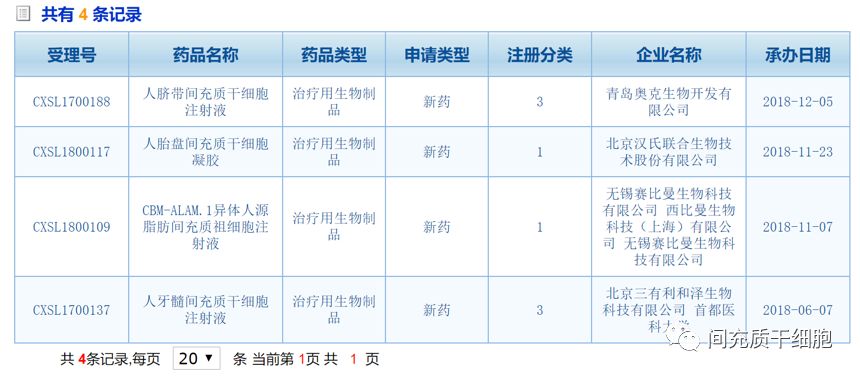
2018年,我国CFDA重新放开细胞产品的注册申请,迈出了历史性的一步。我国MSCs新药注册见下表:
小结:
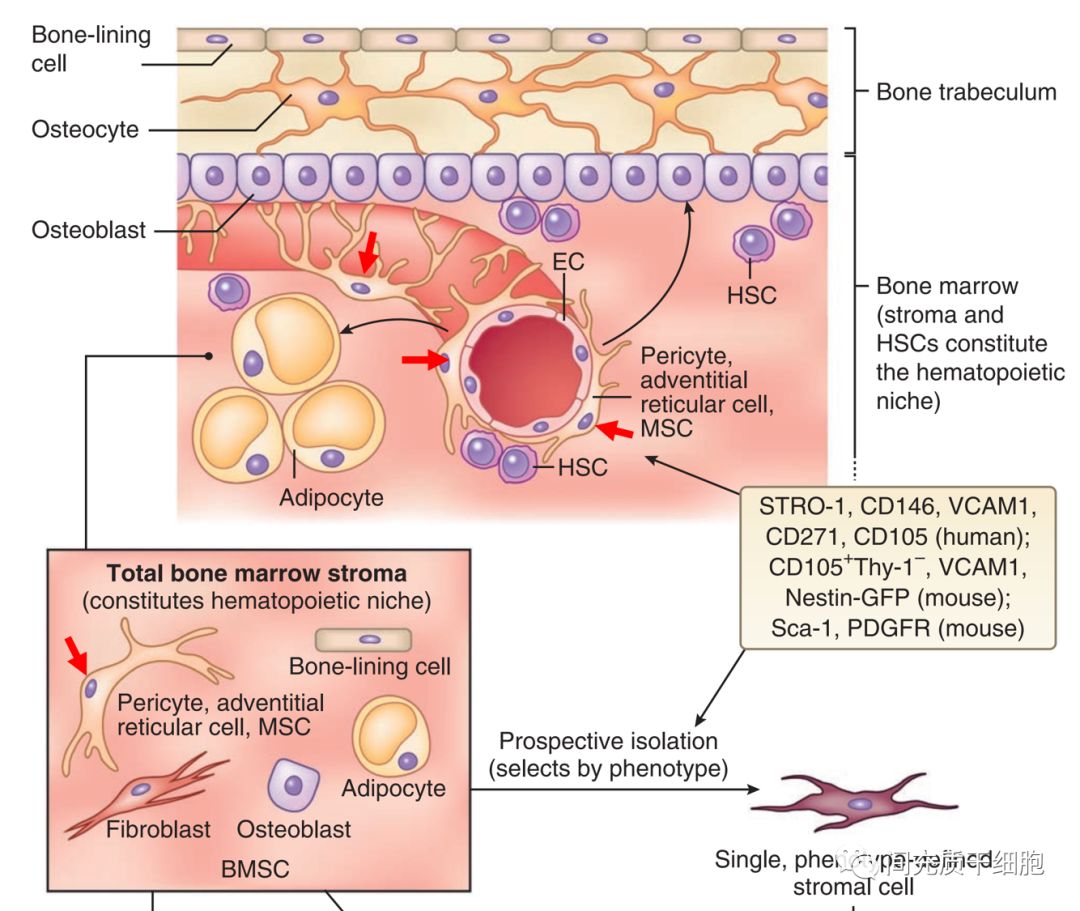
在生理上,MSCs位于血窦的外表面,血窦是骨髓中一种典型的血管类型 (红色箭头所指就是MSCs)。
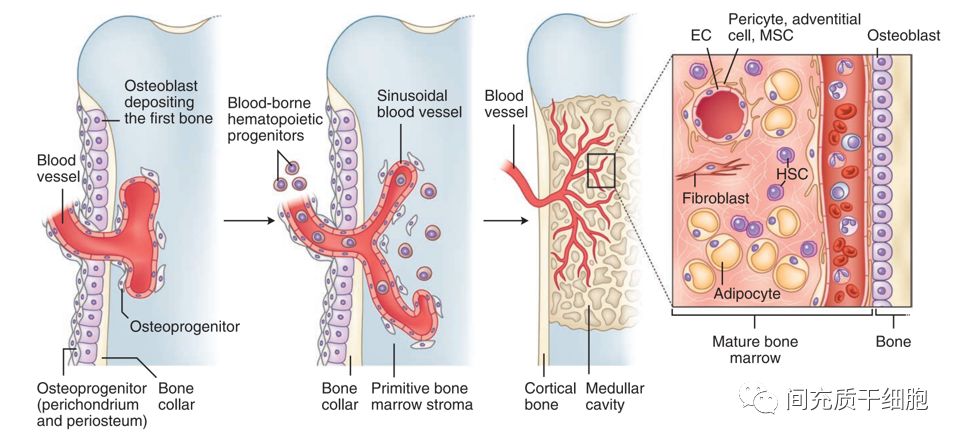
血窦是骨髓中三维血管周围基质间隔的一部分,在骨骼的器官形成过程中起源于原始软骨膜上的成骨细胞,并与生长的血管一起侵入骨底,形成造血干细胞的微环境(niche)(图)。
造血干细胞有着比MSCs更悠久的研究历史,而且在临床应用上,造血干细胞移植后有一部分(约10%)能迁移到骨髓中定居下来,重建了整个造血系统,伴随着整个生命周期。正是由于造血干细胞给人们的印象过于深刻,导致人们也期盼着这个称为“干细胞”的MSCs也像造血干细胞一样,一次成功的输入移植能永久成活。但是,如果深入思考,就会发现MSCs和造血干细胞的功能特性完全不一样,治疗方式也不一样。比如,①造血干细胞移植之前,需要杀死原本身体中存在于骨髓中病变的造血干细胞(俗称清髓),给新来的造血干细胞腾出空间(niche)发挥功能,而现在如何能做到杀死原本身体中存在的MSCs以腾出空间?②造血干细胞只有在自己的巢(niche)中才能长期存活,只有能存活下来才能进行分化,而MSCs的巢在哪里?因此,不能根据造血干细胞的表现来生搬硬套在MSCs身上。
由于MSCs不表达MHC-II类抗原,具有低免疫原性,而且还能抑制免疫反应,因此有专家认为MSCs属于免疫豁免的细胞。但是,大量的动物实验和有限的临床研究均显示输入的异体MSCs均不能在体内长期存在,而且这些数据都是来源于分娩以后的动物和人类。既然MSCs不能在体内长期存在,那么MSCs就无法分化为子代细胞而发挥细胞替代作用,也就是说分化作用不是MSCs的治疗机制。这也是为什么Caplan教授强调MSCs并不是干细胞的原因。即使分化作用不是MSCs的治疗机制,但也不应该否认MSCs的干性(体内自我更新和分化),因为上文已经论述过MSCs在体内具备自我更新和分化的功能。